
生物学はいかに創られたか(9) 最終回:ワトソンとクリックのDNA 二重らせんモデル
ワトソンとクリックのDNA 二重らせんモデル 柴井博四郎(挿絵・今井孝夫)
シャーガフの法則
アベリーに触発されてDNA研究をはじめたシャーガフは、いろいろな生物について、DNAを構成する4塩基の定量分析を行ないました。生物の種類が変わると4塩基の分析値も変化しますが、動物も植物もどんな生物であれ、DNAに含まれるアデニンの分子数とチミンの分子数は等しく、またグアニンとシトシンの分子数も等しいことを明らかにしました。これをシャーガフの法則といいます。
クリックの直感は、この法則がDNAの構造と機能に密接に関連していることに感づいていました。DNA二重らせんモデルはこの法則を満たすものでなければならなかったのです。
DNA二重らせんモデルがワトソンとクリックによって発表されるまでの経過を以下に紹介します。ワトソン著「二重らせんモデル」から引用します。
ワトソンの指導教官ルリアの影響
・そのころルリアはおもに細菌のウイルス(ファージ)の増殖に関する実験をしていた。かなり前からカンがよい遺伝学者たちのあいだでは、ウイルスは裸の遺伝子の一形態ではないだろうかと考えられはじめていた。もしそうだとすれば、遺伝子の実体やその複製のしかたを発見するには、ウイルスの性質を研究するのが手っとり早いことになる。
・ルリアはウイルス(遺伝子)の化学構造が解明されないかぎり真の解答は得られないのではないかと疑っていた。
・(弟子のワトソンを)タンパク質を研究する化学者にするか、核酸の化学者にするかについては、ルリアは迷わなかった。アベリーの実験は、DNAこそ遺伝物質の本体であることをにおわせていた。
モーリスのX線解析像につよい刺激
・ナポリで高分子学会が開かれモーリス・ウイルキンスが講演:「この像は、これまでの写真よりはるかに鮮明な部分まで写し出しており、事実上、結晶物質によって生じたものとみなしてもよいのです。」
・私は化学に対して、はっと目を開かれる思いだった。モーリスの話を聞くまでは、遺伝子とはつかみどころのない不規則なものではなかろうかという心配があった。しかし、いま、遺伝子は結晶しうることを知った。つまり遺伝子とは規則正しい構造をもったものであり、正攻法で解決できるということだ。
・モーリスのことはだんだん忘れてしまったが、彼のDNAの写真は忘れられなかった。生命のナゾを秘めたカギは、私の脳裏に焼きついていた。
ケンブリッジ留学
・私はX線解析像の解読法を学ぶには、どこがよいかを考えはじめた。イギリスのケンブリッジしかない。マックス・ベルツが生体高分子、とくにヘモグロビンタンパクの構造に関心を持っていると聞いていた。そこで私はルリアあてに、自分がかきたてられている新しい情熱について書き送り、なんとかケンブリッジの研究室で受け入れてもらえるようにとりはからう手だてを問い合わせた。
ワトソンとクリックの共同研究
・私はすぐに、フランシス・クリックという愉快な話し相手を見つけた。ベルツの研究室で、タンパク質よりDNAのほうが重要なことを知っている人物に会えたのはまったく幸運だった。
・毎日少なくとも4,5時間は、何にもわずらわされずにクリックと話すことができた。われわれがとくに熱中したのは、どうやってライナス・ポーリングがα―へリックス(タンパク質の立体構造)を発見したかをめぐる、微に入り、細をうがった議論であった。ポーリングの成功のカギは、彼が構造化学の簡単な法則を信頼したことにあった。α―へリックスは、ただX線写真を見つめているうちに発見されたものではなく、決め手となった秘訣は、どの分子がお互いに隣り合っているかを、問うてみることだったのだ。紙と鉛筆のかわりに、子供の玩具そっくりの一組の分子模型であった。だから同じやり方でDNAの構造が解けない理由は何もなかった。やるべきことは分子模型をあれこれいじりまわすことであった。DNAの構造もやはりらせんであろう。
・はじめて話し合ったときから、DNA分子には非常に多数のヌクレオチド分子が規則正しく直線状につながったところがあると仮定した。われわれの推理は、答えは簡単にちがいないという考え方にもとづいていた。
最初のモデル
・モーリスがクリックに話したところによると、DNA分子の直径は、たった1本のポリヌクレオチド鎖だけでは説明できないくらい、大きいということだった。そこで彼は、DNA分子は、数本のポリヌクレオチド鎖がお互いにねじれ合っている複合らせんではないかと考えた。
・塩基の順序は当然、まったく不規則にする必要がある。もし塩基の順序がいつも同じだったら、DNA分子はどれも見分けがつかなくなり、遺伝子のひとつひとつを区別する多様性がなくなってしまう。
・鎖を結びつけている力については、2価のマグネシウムのような陽イオンが、2つかそれ以上のリン酸基を結合させている形の、塩の橋を考えるのが最もよいと思われた。
・その模型は、3本の鎖がらせん状にそって2.8nmごとの結晶学上のくり返しを示すやり方で、お互いにねじれ合っていた。これはモーリスとロザリンド(モーリスの研究室でX線解析像を写している女性)の図にちょうど合う特徴であったから、仕事台から降りて午後の成果を調べなおしているクリックは目に見えて元気になっていた。あと数時間つづければ、かなり立派な模型ができ上がるだろう。
最初のモデルはロザリンドに否定される
・ロザリンドの頭の中では、DNAがらせん構造をとっているという証拠はほんの少しもなかった。ほんとうにらせんかどうかは、X線の研究をさらにつづけて、はじめてわかることだ。
・モデルそのものを見ても彼女の軽べつ感は増しただけだった。マグネシウムイオンは水分子の堅い殻で囲まれているのだから、きちんとした構造の柱にはなれるはずがないと指摘した。
・DNA試料の水分含量についての私の記憶がまちがっていたにちがいないという事実が出てきて、私を当惑させた。正確なDNAモデルは、10倍の水を含んでいなければならない。
・われわれの議論の基礎はまだ甘かったという結論を避けることはできなかった。
クリックの直感
・若い理論化学者、ジョン・グリフィスと何度か話しているうちに、シャーガフの法則は重要なことなのではないかという疑いが、クリックの頭の中でカチリと音を立てた。
・(グリフィスの)ほぼ精密な証明法によって、アデニンとチミンが、その平らな面でお互いにくっつき合っているらしいことがわかった、という計算の結果を聞いた。同様な論法で、グアニンとシトシンの間にも、引力があるといえるだろうということもわかった。
・クリックは、すぐにその答えにとびついた。彼の記憶が正しいとすれば、この塩基対は、シャーガフが等量存在するといっていたものだ。グリフィスの計算とシャーガフの実験結果が一致した。
競争相手ポーリングの動向
・ポーリングは、遅かれ早かれ科学の賞の中での、最高賞を狙って挑戦してくるのは目に見えていた。彼がそれに関心をもっていることは疑いえなかった。ポーリングほどの大化学者が、DNAこそ全分子のなかでも金的を射止めるための最高のものだと気づかぬはずはない。動かしがたい証拠もあった。モーリスはポーリングから結晶DNAのX線写真の複写を一枚送って欲しい、という手紙を受け取っていた。
・ピーター(ポーリングの息子でケンブリッジに留学中)の手にはアメリカからの手紙がにぎられていた。ポーリングがとうとうDNAの構造をつかんだという、われわれが長いあいだ恐れていたニュースが書かれていた。
・ポーリングの(発表前)論文が大西洋を渡ってきた。ケンブリッジに急送されてきた論文は2部で、一部はブラッグ卿(留学先の所長)あて、もう一部はピーターあてになっていた。
ポーリング・モデルの誤りに気づく、時は切迫している
・ポーリング・モデルは中央に糖とリン酸の骨格をもった3本のらせん構造をしていた。それは、途中でお流れになった去年のわれわれの成果とあまりによく似ていた。
・私は一目で、何か誤りがあると感じた。ポーリングの模型では。リン酸基がイオン化しておらず、各基が水素原子を結合しており、したがって全体として電荷がないのである。その水素原子が3本のお互いにねじれ合った鎖を結合させる水素結合にひと役買っていた。リン酸基についた水素原子をとってしまえば、鎖はたちまち開いて、構造は消えてなくなるであろう。
・自分の誤りがわかれば、ポーリングは意地でも正しい構造をつかもうと必死になるであろう。ポーリングがふたたびDNAの追及に専念しだすまで、われわれに与えられた余裕はせいぜい6週間であった。
モーリスから最新情報を受ける
・モーリスは私のことを、誤解をおそれて、うっかり秘密をもらせないよそよそしい知人としてではなく、むしろともに戦う仲間として扱った。驚いたことに、彼は助手のウイルソンを使ってひそかにロザリンドのX線の結果を複写させていることを白状した。
・彼女がDNAに新しい三次元形態があることを示す証拠を得ているというのである。彼らが「B型」構造と呼んでいる新形態を示す写真のプリントをもってきた。
注:室長モーリスと技師ロザリンドの関係は険悪。
二回目のモデル
・私の腹は決まっていた。二本鎖で模型を組み立ててみよう。クリックも賛成してくれるにちがいない。生物学上重要な物質は対になっていることくらいはわかっているだろう。
・翌日、ベルツの部屋に飛び込んでいくと、そこにブラッグ卿がおり、クリックはまだ来ていなかった。私は大急ぎで、B型についてのくわしい説明をひととおりし、DNAはその軸に沿って、3.4nmごとの繰り返しをもつ、らせんであるという証拠としてB型の略図を書いてみせた。ブラッグ卿は少しも反対しなかったばかりか、模型組み立てを成功させるようにと激励してくれた。
注:最初のモデル以後、DNA研究はブラッグ卿に止められていた。
・クリックがとくに重要だと認めたのは、子午線上の0.34nmでの反射が他のどの反射よりも強かったという私の主張であった。この結果から考えられることはただひとつ、プリンとピリミジン塩基が、らせんと垂直の方向に、お互いに0.34nm の間隔で重なり合っているということだった。それに加えて、電子顕微鏡とX線写真の両方から、らせんの直径は約2.0nmであるという確信があった。
・いちばんの障害物は塩基であった。それらが外側に出ているかぎりは問題ないけれども、中へ押しこめるとなると、不規則な塩基配列をもった二本の鎖をどうやってきちんと並べるかというぞっとするような問題が生じてくる。クリックもそのことに関しては、一筋の光明も見いだしていないことを認めざるをえなかった。
・紙にアデニンの輪をぐるぐると描いているうちに突然ひらめいたのだ。DNAの構造のなかでのアデニン残基が純粋のアデニンの結晶で見られると同様の水素結合を作っていると考えた場合、そこに何かしらナゾを解くカギがあるかもしれない。
・DNA分子は同一塩基二個間を水素結合で結び合わせされた、二本のまったく同じ塩基配列順序をもった鎖で構成されていると考えた。
・一本の鎖が、もう一本の鎖が合成される際の鋳型として使われるだろうという推論が容易に成り立つ。遺伝子の複製は、まずはじめに二本の同じ鎖が分離する。それからそれぞれの親の鎖を鋳型にして、二本の新しい娘の鎖が作られ、それによって元の分子とまったく同じ二個のDNA分子ができ上がることになる。
・私はしあわせな気持ちでいっぱいで、ベッドに入っても二時間以上寝つかれず、閉じた目の前では一対のアデニン残基がぐるぐる回転していた。このすばらしい思いつきがまちがっているなどとは、ほとんど考えられなかった。私のたてた案は次の日のお昼までには、もうずたずたに引き裂かれてしまった。
同一の塩基間で結ばれた同一の二本鎖モデルはあっという間に否定される
・アメリカ人のドナヒューがこの構造はうまくいかないと横やりを入れてきた。彼の化学的直感では、グアニンはケト型であるはずで、チミンもエノール型が選ばれているのは誤りで、ケト型の方が好都合だと強調した。
注:当時の教科書ではエノール型がケト型と同じ位置で説明されていた。
・クリックは、この構造がシャーガフの法則をまったく説明していないことも気に入らなかった。
翌朝、正しいモデルに気づく
・翌朝、まだだれも来ていない部屋へ着くと、机の上の紙類をきれいに片づけ、水素結合で結びつけられる塩基の対を並べてみるために、大きく、平らな場所をつくった。
・最初はどうしても同じ塩基同士が結びつくという偏見から抜け出すことができなかった。
・突然、アデニンとチミンの対が二個の水素結合で連結された形は、グアニンとシトシンが最低二個の水素結合で結合された場合とまったく同じであることに気づいた。
・大急ぎでドナヒューを呼び、今度の塩基対について何か異論があるかどうかをたずねた。彼が少しも問題がないといったときには、天にものぼる心地がした。
・なぜプリン残基(アデニンとグアニン)の数が正確にピリミジン残基(チミンとシトシン)の数と等しいのかというナゾが解けたのだ。ここでにわかに、シャーガフの法則がDNA二重らせん構造を示唆するものとして、ぐんとクローズ・アップされてきた。
・私を興奮させたのは、このタイプの二重らせん構造は、前に私が考えていた同じ塩基同士が対を作る場合よりも、はるかに申し分のない複製機構を暗示していることだった。二本のねじれ合った鎖の塩基の配列が決まれば、その相手は自動的に決まってしまう。
・クリックがやってきて、ドアから身体を半分入れるか入れないうちに、私は万事がスッキリする解答が見つかったことを一気に告げた。アデニン—チミンとグアニン—シトシンの組み合わせが、よく似た形をしているという事実には、さすがに大きな衝撃を受けた。
・われわれは昼食に出かけ、こんなに美しい構造は存在しないはずはないと話し合った。
Nature誌の歴史的論文では
・DNA塩の構造について提案したい。この構造は、生物学的につよく興味をひく新しい特徴をもっている。
・核酸の構造は既にポーリングとコリーによって提案されている。その原稿を発表前に私どもに見せて頂いたことを感謝する。彼らのモデルは3本の絡みあった鎖から成り、リン酸が繊維の軸の近くで、塩基は外側に位置する。
・私どもは根源的にことなった構造を提案する。この構造では、二本のらせん鎖が同じ軸を巻いている。
・二本の鎖は軸に対して直角の右まきのらせんで、鎖の中の原子の順序は逆向きに走っている。
・塩基はらせんの内側に、リン酸は外側に位置する。
・らせんに沿ってヌクレオチドが一つ上に進むと、垂直方向に0.34nmのぼり、軸のまわりを36°回転する。したがって、らせんの一巻きは垂直方向3.4nmで10ヌクレオチドが含まれる。
・リン酸は外側にあり、軸の中心からの距離は1nm(らせんの直径は2nm)である。
・この構造が新しい点は、二本の鎖がプリン塩基とピリミジン塩基によって結ばれていることである。塩基は軸に対して直角に位置し、片方の鎖のプリン塩基は他方の鎖のピリミジン塩基と水素結合によって結ばれる。
・生物の中でもっともあり得る塩基の構造が、エノール型ではなくケト型であるとすると、特殊な組み合わせである、アデニンはチミンと、グアニンはシトシンとペアーになる。
・片方の鎖で塩基がならぶ順序は決められないが、片方の鎖の塩基の順序が決まれば、他方の鎖の塩基の順序は自動的に決定されるということを意味している。
・私どもが仮定した特殊な塩基の組み合わせはただちに遺伝子複製機構のありようを暗示している。
・ジュリー・ドナヒューには、原始間の距離について終始助言と批判をいただき感謝する。
・未公表な実験結果とアイデアをもとにしたDNAの一般的な特性に関するモーリス・ウイルキンスとロザリンド・フランクリンの知識から刺激を受けた。
(完)
参考書
ジェームス・ワトソン著 江上不二夫/中村桂子訳「二重らせん」講談社
J. D. Watson and F. H. C. Crick(1953) . “A Structure for Deoxyribose Nucleic Acid”. Nature 171(4356):737-738.
シャーガフの法則
アベリーに触発されてDNA研究をはじめたシャーガフは、いろいろな生物について、DNAを構成する4塩基の定量分析を行ないました。生物の種類が変わると4塩基の分析値も変化しますが、動物も植物もどんな生物であれ、DNAに含まれるアデニンの分子数とチミンの分子数は等しく、またグアニンとシトシンの分子数も等しいことを明らかにしました。これをシャーガフの法則といいます。
クリックの直感は、この法則がDNAの構造と機能に密接に関連していることに感づいていました。DNA二重らせんモデルはこの法則を満たすものでなければならなかったのです。
DNA二重らせんモデルがワトソンとクリックによって発表されるまでの経過を以下に紹介します。ワトソン著「二重らせんモデル」から引用します。
ワトソンの指導教官ルリアの影響
・そのころルリアはおもに細菌のウイルス(ファージ)の増殖に関する実験をしていた。かなり前からカンがよい遺伝学者たちのあいだでは、ウイルスは裸の遺伝子の一形態ではないだろうかと考えられはじめていた。もしそうだとすれば、遺伝子の実体やその複製のしかたを発見するには、ウイルスの性質を研究するのが手っとり早いことになる。
・ルリアはウイルス(遺伝子)の化学構造が解明されないかぎり真の解答は得られないのではないかと疑っていた。
・(弟子のワトソンを)タンパク質を研究する化学者にするか、核酸の化学者にするかについては、ルリアは迷わなかった。アベリーの実験は、DNAこそ遺伝物質の本体であることをにおわせていた。
モーリスのX線解析像につよい刺激
・ナポリで高分子学会が開かれモーリス・ウイルキンスが講演:「この像は、これまでの写真よりはるかに鮮明な部分まで写し出しており、事実上、結晶物質によって生じたものとみなしてもよいのです。」
・私は化学に対して、はっと目を開かれる思いだった。モーリスの話を聞くまでは、遺伝子とはつかみどころのない不規則なものではなかろうかという心配があった。しかし、いま、遺伝子は結晶しうることを知った。つまり遺伝子とは規則正しい構造をもったものであり、正攻法で解決できるということだ。
・モーリスのことはだんだん忘れてしまったが、彼のDNAの写真は忘れられなかった。生命のナゾを秘めたカギは、私の脳裏に焼きついていた。
ケンブリッジ留学
・私はX線解析像の解読法を学ぶには、どこがよいかを考えはじめた。イギリスのケンブリッジしかない。マックス・ベルツが生体高分子、とくにヘモグロビンタンパクの構造に関心を持っていると聞いていた。そこで私はルリアあてに、自分がかきたてられている新しい情熱について書き送り、なんとかケンブリッジの研究室で受け入れてもらえるようにとりはからう手だてを問い合わせた。
ワトソンとクリックの共同研究
・私はすぐに、フランシス・クリックという愉快な話し相手を見つけた。ベルツの研究室で、タンパク質よりDNAのほうが重要なことを知っている人物に会えたのはまったく幸運だった。
・毎日少なくとも4,5時間は、何にもわずらわされずにクリックと話すことができた。われわれがとくに熱中したのは、どうやってライナス・ポーリングがα―へリックス(タンパク質の立体構造)を発見したかをめぐる、微に入り、細をうがった議論であった。ポーリングの成功のカギは、彼が構造化学の簡単な法則を信頼したことにあった。α―へリックスは、ただX線写真を見つめているうちに発見されたものではなく、決め手となった秘訣は、どの分子がお互いに隣り合っているかを、問うてみることだったのだ。紙と鉛筆のかわりに、子供の玩具そっくりの一組の分子模型であった。だから同じやり方でDNAの構造が解けない理由は何もなかった。やるべきことは分子模型をあれこれいじりまわすことであった。DNAの構造もやはりらせんであろう。
・はじめて話し合ったときから、DNA分子には非常に多数のヌクレオチド分子が規則正しく直線状につながったところがあると仮定した。われわれの推理は、答えは簡単にちがいないという考え方にもとづいていた。
最初のモデル
・モーリスがクリックに話したところによると、DNA分子の直径は、たった1本のポリヌクレオチド鎖だけでは説明できないくらい、大きいということだった。そこで彼は、DNA分子は、数本のポリヌクレオチド鎖がお互いにねじれ合っている複合らせんではないかと考えた。
・塩基の順序は当然、まったく不規則にする必要がある。もし塩基の順序がいつも同じだったら、DNA分子はどれも見分けがつかなくなり、遺伝子のひとつひとつを区別する多様性がなくなってしまう。
・鎖を結びつけている力については、2価のマグネシウムのような陽イオンが、2つかそれ以上のリン酸基を結合させている形の、塩の橋を考えるのが最もよいと思われた。
・その模型は、3本の鎖がらせん状にそって2.8nmごとの結晶学上のくり返しを示すやり方で、お互いにねじれ合っていた。これはモーリスとロザリンド(モーリスの研究室でX線解析像を写している女性)の図にちょうど合う特徴であったから、仕事台から降りて午後の成果を調べなおしているクリックは目に見えて元気になっていた。あと数時間つづければ、かなり立派な模型ができ上がるだろう。
最初のモデルはロザリンドに否定される
・ロザリンドの頭の中では、DNAがらせん構造をとっているという証拠はほんの少しもなかった。ほんとうにらせんかどうかは、X線の研究をさらにつづけて、はじめてわかることだ。
・モデルそのものを見ても彼女の軽べつ感は増しただけだった。マグネシウムイオンは水分子の堅い殻で囲まれているのだから、きちんとした構造の柱にはなれるはずがないと指摘した。
・DNA試料の水分含量についての私の記憶がまちがっていたにちがいないという事実が出てきて、私を当惑させた。正確なDNAモデルは、10倍の水を含んでいなければならない。
・われわれの議論の基礎はまだ甘かったという結論を避けることはできなかった。
クリックの直感
・若い理論化学者、ジョン・グリフィスと何度か話しているうちに、シャーガフの法則は重要なことなのではないかという疑いが、クリックの頭の中でカチリと音を立てた。
・(グリフィスの)ほぼ精密な証明法によって、アデニンとチミンが、その平らな面でお互いにくっつき合っているらしいことがわかった、という計算の結果を聞いた。同様な論法で、グアニンとシトシンの間にも、引力があるといえるだろうということもわかった。
・クリックは、すぐにその答えにとびついた。彼の記憶が正しいとすれば、この塩基対は、シャーガフが等量存在するといっていたものだ。グリフィスの計算とシャーガフの実験結果が一致した。
競争相手ポーリングの動向
・ポーリングは、遅かれ早かれ科学の賞の中での、最高賞を狙って挑戦してくるのは目に見えていた。彼がそれに関心をもっていることは疑いえなかった。ポーリングほどの大化学者が、DNAこそ全分子のなかでも金的を射止めるための最高のものだと気づかぬはずはない。動かしがたい証拠もあった。モーリスはポーリングから結晶DNAのX線写真の複写を一枚送って欲しい、という手紙を受け取っていた。
・ピーター(ポーリングの息子でケンブリッジに留学中)の手にはアメリカからの手紙がにぎられていた。ポーリングがとうとうDNAの構造をつかんだという、われわれが長いあいだ恐れていたニュースが書かれていた。
・ポーリングの(発表前)論文が大西洋を渡ってきた。ケンブリッジに急送されてきた論文は2部で、一部はブラッグ卿(留学先の所長)あて、もう一部はピーターあてになっていた。
ポーリング・モデルの誤りに気づく、時は切迫している
・ポーリング・モデルは中央に糖とリン酸の骨格をもった3本のらせん構造をしていた。それは、途中でお流れになった去年のわれわれの成果とあまりによく似ていた。
・私は一目で、何か誤りがあると感じた。ポーリングの模型では。リン酸基がイオン化しておらず、各基が水素原子を結合しており、したがって全体として電荷がないのである。その水素原子が3本のお互いにねじれ合った鎖を結合させる水素結合にひと役買っていた。リン酸基についた水素原子をとってしまえば、鎖はたちまち開いて、構造は消えてなくなるであろう。
・自分の誤りがわかれば、ポーリングは意地でも正しい構造をつかもうと必死になるであろう。ポーリングがふたたびDNAの追及に専念しだすまで、われわれに与えられた余裕はせいぜい6週間であった。
モーリスから最新情報を受ける
・モーリスは私のことを、誤解をおそれて、うっかり秘密をもらせないよそよそしい知人としてではなく、むしろともに戦う仲間として扱った。驚いたことに、彼は助手のウイルソンを使ってひそかにロザリンドのX線の結果を複写させていることを白状した。
・彼女がDNAに新しい三次元形態があることを示す証拠を得ているというのである。彼らが「B型」構造と呼んでいる新形態を示す写真のプリントをもってきた。
注:室長モーリスと技師ロザリンドの関係は険悪。
二回目のモデル
・私の腹は決まっていた。二本鎖で模型を組み立ててみよう。クリックも賛成してくれるにちがいない。生物学上重要な物質は対になっていることくらいはわかっているだろう。
・翌日、ベルツの部屋に飛び込んでいくと、そこにブラッグ卿がおり、クリックはまだ来ていなかった。私は大急ぎで、B型についてのくわしい説明をひととおりし、DNAはその軸に沿って、3.4nmごとの繰り返しをもつ、らせんであるという証拠としてB型の略図を書いてみせた。ブラッグ卿は少しも反対しなかったばかりか、模型組み立てを成功させるようにと激励してくれた。
注:最初のモデル以後、DNA研究はブラッグ卿に止められていた。
・クリックがとくに重要だと認めたのは、子午線上の0.34nmでの反射が他のどの反射よりも強かったという私の主張であった。この結果から考えられることはただひとつ、プリンとピリミジン塩基が、らせんと垂直の方向に、お互いに0.34nm の間隔で重なり合っているということだった。それに加えて、電子顕微鏡とX線写真の両方から、らせんの直径は約2.0nmであるという確信があった。
・いちばんの障害物は塩基であった。それらが外側に出ているかぎりは問題ないけれども、中へ押しこめるとなると、不規則な塩基配列をもった二本の鎖をどうやってきちんと並べるかというぞっとするような問題が生じてくる。クリックもそのことに関しては、一筋の光明も見いだしていないことを認めざるをえなかった。
・紙にアデニンの輪をぐるぐると描いているうちに突然ひらめいたのだ。DNAの構造のなかでのアデニン残基が純粋のアデニンの結晶で見られると同様の水素結合を作っていると考えた場合、そこに何かしらナゾを解くカギがあるかもしれない。
・DNA分子は同一塩基二個間を水素結合で結び合わせされた、二本のまったく同じ塩基配列順序をもった鎖で構成されていると考えた。
・一本の鎖が、もう一本の鎖が合成される際の鋳型として使われるだろうという推論が容易に成り立つ。遺伝子の複製は、まずはじめに二本の同じ鎖が分離する。それからそれぞれの親の鎖を鋳型にして、二本の新しい娘の鎖が作られ、それによって元の分子とまったく同じ二個のDNA分子ができ上がることになる。
・私はしあわせな気持ちでいっぱいで、ベッドに入っても二時間以上寝つかれず、閉じた目の前では一対のアデニン残基がぐるぐる回転していた。このすばらしい思いつきがまちがっているなどとは、ほとんど考えられなかった。私のたてた案は次の日のお昼までには、もうずたずたに引き裂かれてしまった。
同一の塩基間で結ばれた同一の二本鎖モデルはあっという間に否定される
・アメリカ人のドナヒューがこの構造はうまくいかないと横やりを入れてきた。彼の化学的直感では、グアニンはケト型であるはずで、チミンもエノール型が選ばれているのは誤りで、ケト型の方が好都合だと強調した。
注:当時の教科書ではエノール型がケト型と同じ位置で説明されていた。
・クリックは、この構造がシャーガフの法則をまったく説明していないことも気に入らなかった。
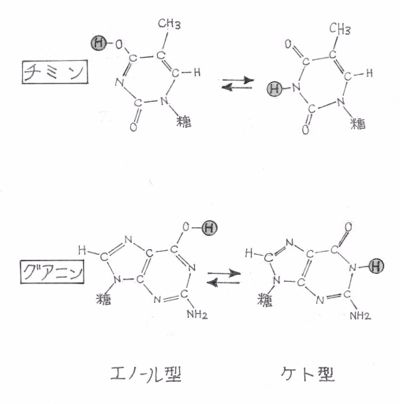 |
| グアニンとチミンの互変異性型 位置を変えうる水素は太字で表している。 |
翌朝、正しいモデルに気づく
・翌朝、まだだれも来ていない部屋へ着くと、机の上の紙類をきれいに片づけ、水素結合で結びつけられる塩基の対を並べてみるために、大きく、平らな場所をつくった。
・最初はどうしても同じ塩基同士が結びつくという偏見から抜け出すことができなかった。
・突然、アデニンとチミンの対が二個の水素結合で連結された形は、グアニンとシトシンが最低二個の水素結合で結合された場合とまったく同じであることに気づいた。
・大急ぎでドナヒューを呼び、今度の塩基対について何か異論があるかどうかをたずねた。彼が少しも問題がないといったときには、天にものぼる心地がした。
・なぜプリン残基(アデニンとグアニン)の数が正確にピリミジン残基(チミンとシトシン)の数と等しいのかというナゾが解けたのだ。ここでにわかに、シャーガフの法則がDNA二重らせん構造を示唆するものとして、ぐんとクローズ・アップされてきた。
・私を興奮させたのは、このタイプの二重らせん構造は、前に私が考えていた同じ塩基同士が対を作る場合よりも、はるかに申し分のない複製機構を暗示していることだった。二本のねじれ合った鎖の塩基の配列が決まれば、その相手は自動的に決まってしまう。
・クリックがやってきて、ドアから身体を半分入れるか入れないうちに、私は万事がスッキリする解答が見つかったことを一気に告げた。アデニン—チミンとグアニン—シトシンの組み合わせが、よく似た形をしているという事実には、さすがに大きな衝撃を受けた。
・われわれは昼食に出かけ、こんなに美しい構造は存在しないはずはないと話し合った。
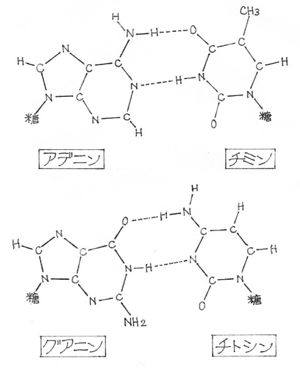 |
| アデニンとチミン、グアニンとシトシンの塩基対 (現在では、グアニンとシトシンのあいだには3本の水素結合があることが分かっている。) |
Nature誌の歴史的論文では
・DNA塩の構造について提案したい。この構造は、生物学的につよく興味をひく新しい特徴をもっている。
・核酸の構造は既にポーリングとコリーによって提案されている。その原稿を発表前に私どもに見せて頂いたことを感謝する。彼らのモデルは3本の絡みあった鎖から成り、リン酸が繊維の軸の近くで、塩基は外側に位置する。
・私どもは根源的にことなった構造を提案する。この構造では、二本のらせん鎖が同じ軸を巻いている。
・二本の鎖は軸に対して直角の右まきのらせんで、鎖の中の原子の順序は逆向きに走っている。
・塩基はらせんの内側に、リン酸は外側に位置する。
・らせんに沿ってヌクレオチドが一つ上に進むと、垂直方向に0.34nmのぼり、軸のまわりを36°回転する。したがって、らせんの一巻きは垂直方向3.4nmで10ヌクレオチドが含まれる。
・リン酸は外側にあり、軸の中心からの距離は1nm(らせんの直径は2nm)である。
・この構造が新しい点は、二本の鎖がプリン塩基とピリミジン塩基によって結ばれていることである。塩基は軸に対して直角に位置し、片方の鎖のプリン塩基は他方の鎖のピリミジン塩基と水素結合によって結ばれる。
・生物の中でもっともあり得る塩基の構造が、エノール型ではなくケト型であるとすると、特殊な組み合わせである、アデニンはチミンと、グアニンはシトシンとペアーになる。
・片方の鎖で塩基がならぶ順序は決められないが、片方の鎖の塩基の順序が決まれば、他方の鎖の塩基の順序は自動的に決定されるということを意味している。
・私どもが仮定した特殊な塩基の組み合わせはただちに遺伝子複製機構のありようを暗示している。
・ジュリー・ドナヒューには、原始間の距離について終始助言と批判をいただき感謝する。
・未公表な実験結果とアイデアをもとにしたDNAの一般的な特性に関するモーリス・ウイルキンスとロザリンド・フランクリンの知識から刺激を受けた。
 |
| DNAの二重らせんモデル |
(完)
参考書
ジェームス・ワトソン著 江上不二夫/中村桂子訳「二重らせん」講談社
J. D. Watson and F. H. C. Crick(1953) . “A Structure for Deoxyribose Nucleic Acid”. Nature 171(4356):737-738.

ご意見、コメントがありましたら、「ユーザ登録」(新規)の上、「ログイン」して「このトピックに投稿する」で書き込み下さい。
登録済み方は、ID(メールアドレス)、パスワードで「ログイン」出来ます。
参照のために4MB以下のファイル添付や本文中にURLの提示も可能です。
なお、投稿者は登録時の「ニックネーム」で表示されます。
投稿内容は、このサイトで公開されると共に、本アイテムの過去投稿者全員にメール連絡されますので、ご承知下さい。
また、非公開をご希望の場合は、トップページ右横にある「問合せ・ご意見」からお知らせ下さい。